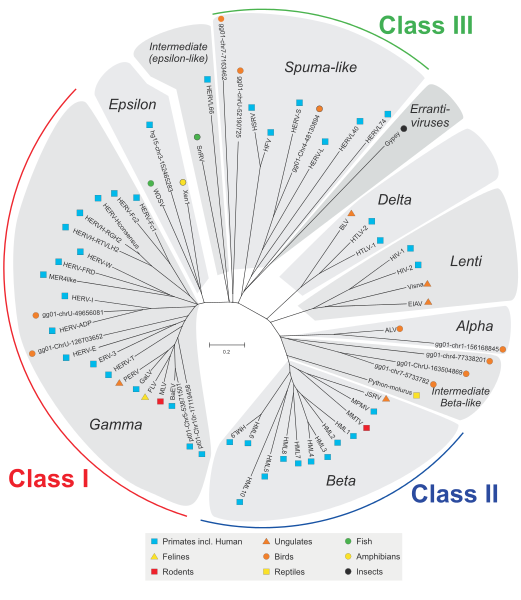
Endogenous retroviruses (ERVs) are endogenous viral elements in the genome that closely resemble and can be derived from retroviruses. They are abundant in the genomes of jawed vertebrates, and they comprise up to 5–8% of the human genome (lower estimates of ~1%).[1][2] ERVs are a subclass of a type of gene called a transposon, which can be packaged and moved within the genome to serve a vital role in gene expression and in regulation.[3][4] Researchers have suggested that retroviruses evolved from a type of transposable gene called a retrotransposon, which includes ERVs; these genes can mutate and instead of moving to another location in the genome they can become exogenous or pathogenic. This means that not all ERVs may have originated as an insertion by a retrovirus but that some may have been the source for the genetic information in the retroviruses they resemble
Terjemahan :
Retrovirus endogen (ERVs) adalah unsur virus endogen dalam genom yang sangat mirip dan dapat diturunkan dari retrovirus . Mereka berlimpah dalam genom vertebrata berahang , dan mereka terdiri sampai 5-8% dari genom manusia (perkiraan lebih rendah dari ~ 1%). [1] [2] ERVs adalah subclass dari jenis gen yang disebut transposon , yang dapat dikemas dan dipindahkan dalam genom untuk melayani peran penting dalam ekspresi gen dan regulasi . [3] [4] para peneliti telah menyarankan bahwa retrovirus berevolusi dari jenis gen transposabel disebut retrotransposon , yang meliputi ERVs; gen ini dapat bermutasi dan bukannya pindah ke lokasi lain dalam genom mereka dapat menjadi eksogen atau patogen. Ini berarti bahwa tidak semua ERVs mungkin berasal sebagai penyisipan oleh retrovirus tetapi beberapa mungkin telah menjadi sumber untuk informasi genetik dalam retrovirus mereka menyerupai.
Peran dalam evolusi genom
retrovirus endogen dapat berperan aktif dalam membentuk genom. Kebanyakan penelitian di daerah ini telah berfokus pada genom manusia dan primata yang lebih tinggi, tetapi vertebrata lainnya, seperti tikus dan domba, juga telah dipelajari secara mendalam. [6] [7] [8] [9] Terminal panjang berulang ( LTR ) urutan yang mengapit genom ERV sering bertindak sebagai alternatif promotor dan enhancer , sering berkontribusi terhadap transcriptome dengan memproduksi varian spesifik jaringan. Selain itu, retroviral protein sendiri telah dikooptasi untuk melayani fungsi tuan rumah baru, khususnya dalam reproduksi dan pengembangan. Rekombinasi antara urutan retroviral homolog juga berkontribusi pertukaran gen dan generasi variasi genetik. Selanjutnya, dalam contoh dari efek berpotensi antagonis urutan retroviral, gen penekan memiliki co-berevolusi untuk memerangi mereka.
Ltrs Solo dan ltrs terkait dengan urutan retroviral lengkap telah ditunjukkan untuk bertindak sebagai elemen transkripsi gen tuan rumah. Berbagai aksi mereka terutama oleh penyisipan ke dalam ‘5 UTRs gen protein coding; Namun, mereka telah dikenal untuk bertindak atas gen hingga 70-100 kb pergi. [6] [10] [11] [12] Mayoritas elemen ini dimasukkan ke arah akal untuk gen yang sesuai mereka, tetapi telah ada bukti ltrs bertindak dalam arah antisense dan sebagai promotor dua arah untuk gen tetangga. [13] [14] dalam beberapa kasus, fungsi LTR sebagai promotor utama untuk gen. Misalnya, pada manusia AMY1C memiliki urutan ERV lengkap di daerah promotor nya; yang terkait LTR menganugerahkan ekspresi tertentu ludah dari pencernaan enzim amilase . [15] Juga, promotor utama untuk empedu asam-CoA: asam amino N-acyltransferase (Baat), yang kode untuk enzim yang tidak terpisahkan dalam metabolisme empedu, adalah LTR asal. [11] [16]
Penyisipan solo ERV-9 LTR mungkin telah menghasilkan fungsional open reading frame (ORF), menyebabkan kelahiran kembali manusia kekebalan terkait gen GTPase (IRGM). [17] sisipan ERV juga telah ditunjukkan untuk menghasilkan situs sambatan alternatif baik dengan integrasi langsung ke gen, seperti dengan leptin manusia reseptor hormon, atau didorong oleh ekspresi dari LTR hulu, seperti dengan fosfolipase A-2 seperti protein. [18]
Sebagian besar waktu, namun, fungsi LTR sebagai salah satu dari banyak promotor alternatif, sering berunding ekspresi jaringan-spesifik yang berhubungan dengan reproduksi dan pengembangan. Bahkan, 64% dari varian transkripsi diketahui LTR-dipromosikan yang dinyatakan dalam jaringan reproduksi. [19] Sebagai contoh, kode gen CYP19 untuk aromatase P450, enzim penting untuk sintesis estrogen, yang biasanya dinyatakan dalam otak dan organ reproduksi kebanyakan mamalia. [11] Namun, pada primata, varian transkripsi LTR-dipromosikan menganugerahkan ekspresi plasenta dan bertanggung jawab untuk mengontrol kadar estrogen selama kehamilan. [11] Selanjutnya, apoptosis neuronal penghambatan protein (naip), biasanya luas, memiliki sebuah LTR dari keluarga HERV-P bertindak sebagai promotor yang menganugerahkan ekspresi ke testis dan prostat. [20] protein lain, seperti asam nitrat sintase 3 (NOS3), interleukin-2 B reseptor (IL2RB), dan mediator lain sintesis estrogen, HSD17B1, juga alternatif diatur oleh ltrs yang memberi ekspresi plasenta, namun fungsi spesifik mereka belum diketahui. [16] [21] tingginya tingkat ekspresi reproduksi diduga menjadi efek lanjutan dari metode yang mereka endogenized; Namun, ini juga mungkin karena kurangnya metilasi DNA pada jaringan kuman-line. [16]
Misalnya yang terbaik-ditandai ekspresi protein plasenta tidak berasal dari gen tuan rumah alternatif dipromosikan tetapi dari kooptasi lengkap dari protein retroviral. Retroviral protein env fusogenik, yang berperan dalam masuknya virion ke dalam sel inang, memiliki dampak penting pada perkembangan plasenta mamalia. Pada mamalia, protein env utuh disebut syncytins bertanggung jawab untuk pembentukan dan fungsi syncytiotrofoblas . [8] Sel-sel berinti terutama bertanggung jawab untuk menjaga pertukaran nutrisi dan memisahkan janin dari sistem kekebalan tubuh ibu. [8] Ia telah mengemukakan bahwa seleksi dan fiksasi protein ini untuk fungsi ini telah memainkan peran penting dalam evolusi viviparity . [22]
Selain itu, masuknya ERVs dan ltrs masing-masing memiliki potensi untuk menginduksi kromosom penataan ulang karena rekombinasi antara urutan virus pada lokus antar kromosom. Penataan ulang ini telah terbukti menyebabkan duplikasi gen dan penghapusan yang sebagian besar berkontribusi genom plastisitas dan secara dramatis mengubah dinamika fungsi gen. [23] Selanjutnya, retroelements secara umum sebagian besar terjadi di berkembang pesat, keluarga gen mamalia khusus yang fungsinya sebagian besar terkait dengan respon terhadap stres dan rangsangan eksternal. [11] Secara khusus, baik manusia kelas I dan kelas II MHC gen memiliki kepadatan tinggi elemen HERV dibandingkan dengan keluarga multi-lokus-gen lainnya. [18] Hal ini telah ditunjukkan yang HERVs telah memberi kontribusi pada pembentukan ekstensif digandakan duplicon blok yang membentuk HLA kelas 1 keluarga gen. [24] Lebih khusus, HERVs terutama menempati daerah di dalam dan di antara break point antara blok ini, menunjukkan bahwa cukup duplikasi dan penghapusan acara , biasanya terkait dengan crossover yang tidak sama, memfasilitasi pembentukan mereka. [25] generasi dari blok ini, diwariskan sebagai immunohaplotypes, bertindak sebagai polimorfisme pelindung terhadap berbagai antigen yang mungkin telah dijiwai manusia dengan keuntungan lebih dari primata lainnya. [24]
Akhirnya, penyisipan ERVs atau elemen ERV ke daerah genic DNA host, atau berlebih dari varian transkripsi mereka, memiliki potensi yang jauh lebih tinggi untuk menghasilkan efek merusak dari yang positif. Penampilan mereka ke dalam genom telah menciptakan co-evolusi dinamis host-parasit yang berkembang biak duplikasi dan perluasan gen represor. Contoh yang paling jelas dari ini melibatkan duplikasi cepat dan proliferasi tandem zinc-finger gen dalam genom mamalia. Gen seng-jari, terutama mereka yang termasuk domain KRAB, ada di nomor copy tinggi dalam genom vertebrata, dan jangkauan mereka fungsi terbatas pada transkripsi peran. [26] Hal ini telah ditunjukkan pada mamalia, bagaimanapun, bahwa diversifikasi ini gen adalah karena beberapa duplikasi dan fiksasi peristiwa dalam menanggapi urutan retroviral baru atau salinan endogen mereka untuk menekan transkripsi mereka.
Sumber : #en.wikipedia.org.

